39 Null models aren’t dull models
One of the fun things about science is discovering concepts that are not intuitive, at least to some of us, but are nonetheless important. Evolution by natural selection is certainly the most powerful of these. For me, I would add null models, compound interest, marginal value theorem, and genetic drift.
Here I will address only null models. When I started in biology in the late 1960s, much of the ecological literature focused on describing patterns observed in nature. One might compare microhabitat associations of two species in the same habitat: if those species differed in microhabitat usage (either all or in part), ecologists typically concluded that this difference reduced competition for space and perhaps for food. The general protocol was to observe a pattern and then infer a process that presumably generated this pattern.
Here is a behavioral example. Ever since Cowles and Bogert (1944) drew attention to “behavioral thermoregulation,” biologists found that many desert reptiles (and many other ectotherms) adjust their behavior to control their body temperature. Early or late in the day, when environmental temperatures are cool, reptiles bask in the sun, thus raising body temperatures above air temperatures. In the middle of the day, when environmental temperatures are hot and potentially lethal, reptiles stay in the shade or move to a cool retreat and only occasionally venture out into the midday sun. This shuttling between sun and shade enables remarkable control over body temperature.
The concept that behavior could control physiological rates was exciting, especially for herpetologists. Consequently, many herpetologists headed to the field with special fast-reading thermometers (“Schultheis”) and recorded the body temperature of every reptile they caught. This was easy to do and generated much data for minimal effort. Moreover, herpetologists soon realized that the body temperatures selected by individual species influenced the times of day it could be active and what types of habitats it could tolerate. Thus body temperature was ecologically relevant.
When Eric Pianka, Larry Coons, and I went to the Kalahari in 1969, we were fully equipped with Schultheis thermometers. Eric had previously taken thousands of temperatures of desert lizards in North America and in Australia, and he had documented interspecific and geographic variation in those temperatures.
We soon learned that environmental temperatures – and less so competition and predation – drove the behavior of Kalahari lizards. In summer, for example, Trachylepis striata, a less than attractive skink that is active on tree trunks, would usually bask on the sunny side of a tree in the early morning and again late afternoon, but would be in the shade at midday. In late autumn and winter, these lizards were active only at midday and were almost always in the sun. This pattern is exactly what one would predict, if the lizards were thermoregulating. Observe pattern then infer process.
I was happy with my interpretation of these patterns until we were visited by Wulf Haacke (herpetologist at the Transvaal Museum) and a botanist, whose name I have long forgotten. I described the behavioral shifts I’d seen in response to temperature and seasons and attributed them to thermoregulation by the lizards.
The botanist asked for following question: how did I know that the lizards don’t thermoregulate but are merely follow insects, which were thermoregulating?
I was somewhat offended by the implication that insects can thermoregulate, but lizards cannot. I countered that lizards thermoregulate carefully in a temperature gradient without any food present; thus, they can no doubt do thermoregulate in nature as well. Even so, the botanist’s question started me thinking about the kinds of robust evidence required to infer process from pattern.
The longer I stayed in the Kalahari, the stronger my view that the behavior and ecology of Kalahari lizards was driven by temperature. I eventually decided to do my Ph.D. thesis on the ecology of behavioral thermoregulation. Null hypotheses soon became a part of my thinking about thermoregulation.
39.1 Heath’s beer can experiment
As a grad student, I read a paper by James Edward Heath (1964) in Science. That paper changed the way I thought about biology in general and thermoregulation in particular.
Before explaining Heath’s paper and its significance, I must put that paper into its historical context. This short paper appeared in an era when herpetologists were grabbing and jabbing every lizard they could catch, and then stating that the resulting data were evidence of thermoregulation. A typical histogram of body temperature is shown below (left panel). These distributions were not symmetrical but highly skewed and truncated at the high end. The long left tail of the distribution was interpreted as coming from lizards that were warming up early in the day, and the short right tail was interpreted as evidence that lizards were behaviorally avoiding high (potentially stressful or even lethal) temperatures. Ergo, pattern was used to predict process.
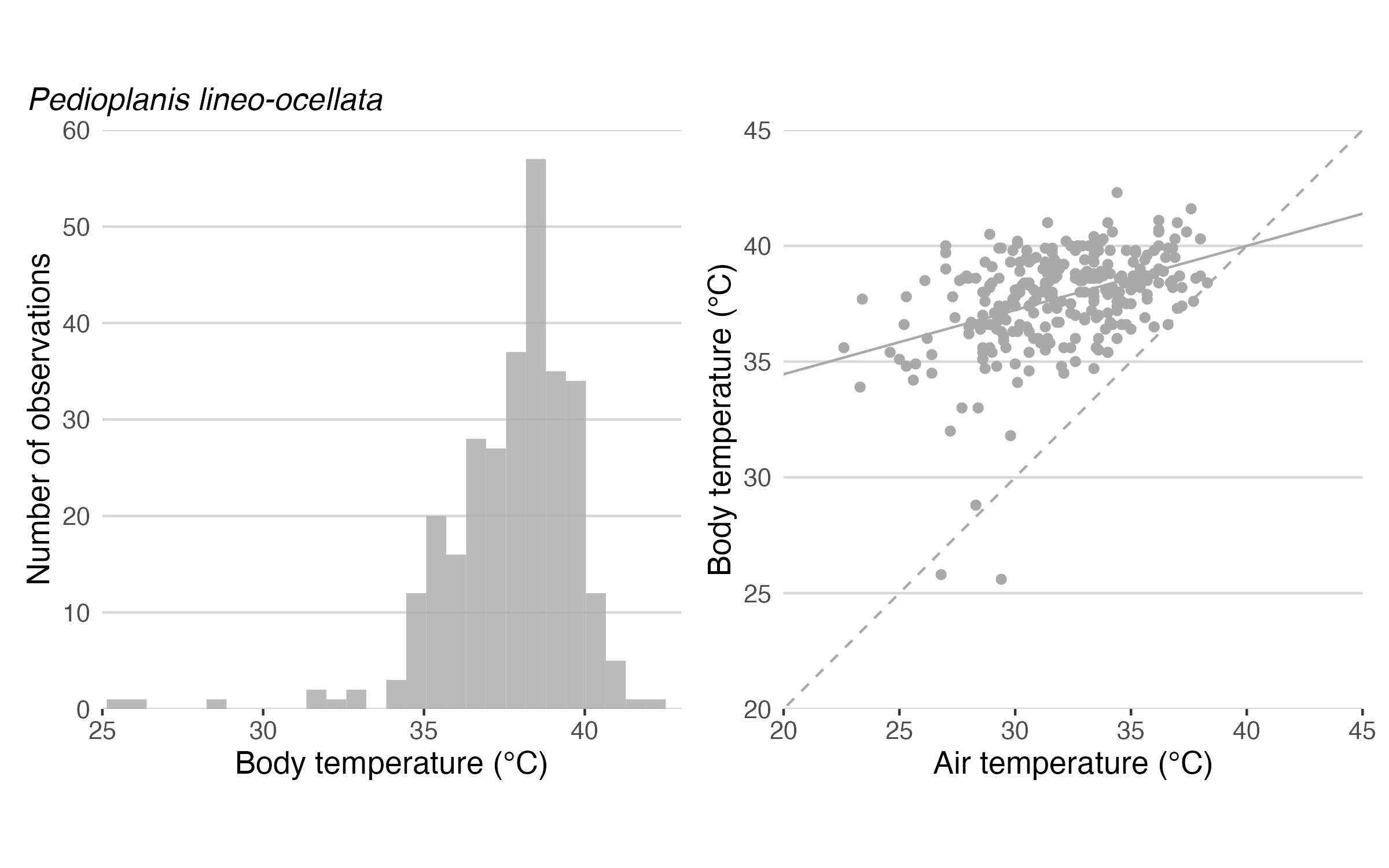
In addition, many authors would plot body temperature against simultaneous air temperature. Often the slope was less than 1; thus, body temperature was elevated well above air temperatures when air temperatures were cool, but converged on air temperatures when air temperatures were hot (see right panel, above). This pattern was interpreted to imply that lizards adjusted the amount of time they spent in sun and shade (mainly in sun early and late, mainly in shade midday). Such adjustments enabled lizards to be warm even early and late in the day as well as to reduce variations in body temperature during the day. Again, pattern was used to predict process.
Herpetologists were not alone in thinking that pattern predicts process. In fact, much of contemporary ecology at was dominated by such thinking. However, Robert Treat Paine challenged that view by arguing that the only way to validate the involvement of a process was to do a manipulative experiment. Bob’s experimental manipulations in the intertidal remain classic examples of why patterns can be misleading, and of why controlled experimental manipulations are necessary to determine the processes behind patterns.
But let’s return to Jim Heath. For his thesis research he conducted a series of elegant and clever laboratory experiments on thermoregulatory behavior of horned lizards. His monograph is great fun to read and is a classic in experimental biology in the lab (Heath 1965). Heath knew that pattern need not imply process and challenged contemporary paradigms in field ecology.
Heath conduced a field experiment demonstrating dramatically that observed patterns can be misleading, and he did so with style. Heath went to a desert and put out 13 Schlitz beer cans, filled with water. He then recorded the “body” temperature of the beer cans over the course of the day, and also air temperatures measured at the same time.
When Heath plotted a histogram of body temperatures of these “inanimate” beer cans, the distribution was remarkably like that of live lizards (see figure above): left skewed, and truncated at high temperatures. This was the pattern expected for a thermoregulating lizard, but Heath’s beer cans were inanimate and not doing anything!
Next he plotted beer can temperature vs. air temperature, and the slope was less than 1, as is often seen with real lizards (right panel, above). Thus, beer cans were relatively warm early in the day, but converged on air temperature at midday. This pattern was expected for a thermoregulating lizard, but not for an inanimate beer can. How could this be?
Heath’s paper attracted attention in part it was published in Science but mainly because Heath used beer cans. That was bold. However, many herpetologists were initially confused by the results and thought that Heath was claiming that lizards do not thermoregulate. He was not saying that at all: he was saying that a histogram of body temperatures, or a regression of Tb on Ta, does not necessarily imply any active thermoregulation by lizards.
Heath argued that what one needed was a control – something that enabled temperatures of lizard-like objects that did not thermoregulate. He suggested a lizard tethered in the field would work. The observed difference between the body temperature of a free-ranging lizard and that of a tethered lizard, could partly be due to thermoregulation.
As a young grad student, I struggled with Heath’s paper. I ‘sort of got it, but sort of didn’t.’ The reason was that Heath had introduced me to a concept that was new to me and to much of ecology, that of a “null model.” The idea here is to ask, not what is the expected pattern for an animal doing something (thermoregulating) but rather to ask, what is the expected pattern for an animal moving randomly or not doing something, or in our case, not thermoregulating. Compare the real and null patterns to determine whether the animal appears to be behaving randomly or not.
[The word “null” was adopted from statistics, where the null hypothesis is an expectation of no effect of some variable or of no difference between two species. That null is central to standard statistical testing of hypothesis.]
This idea is important. Classical studies in ecology looked to see if a pattern matched that expected for some process, and if so, this was taken as support for that process causing the pattern. Heath flipped that on its head, and quantified the pattern expected if the process (thermoregulation) were not involved.
Heath’s beer cans were the first (to my knowledge) empirical null models in ecology. Two decades later, nul models had become a key tool of contemporary ecology and evolution.
Around 1977, Eric Pianka and I were writing a paper on seasonal patterns of thermoregulation of Kalahari lizards. Our data were descriptive – thus, they were patterns. However, we asked whether our observed patterns were sufficient to infer process. We included a special section and an appendix (see below) in our paper that addressed this question.
We needed to discuss Heath’s experiment. When I reread his paper (and some subsequent commentaries in Science), I realized that his beer cans were not really inanimate. For one thing the beer cans were out only from 10 am to 3 pm; such regulation of activity time is perhaps the most common behavioral adjustment that lizards use to limit variation in body temperature. Moreover, if a beer can became shaded, Heath moved it back into the sun; thus Heath controlled basking behavior. But I brooded about the regression of Tb on Ta: Why was it less than 1? This implied that the beer cans were warmer than air temperature early in the day. Why would that happen?
Suddenly I understood. Heath’s beer cans were standing upright – they had to be or the water would have drained. However, this means that early in the day, when the sun was low on the horizon, the long axis of the beer can would have been oriented perpendicular to the rays of the sun. A large fraction of the can’s surface area would be directly exposed to solar radiation, and result in elevated temperatures. At midday, when the sun was high in the sky, the top of the beer can, with its small surface area, would be exposed to the sun and so heating would be reduced, especially if the top was highly reflective (Warren Porter, pers. comm.). This change in orientation relative to the sun’s rays is what real lizards do. [Heath’s 1965 monograph provides a beautiful example.]
Our conclusion that Heath’s beer cans were not 100% inanimate does not negate his assertions. Instead, it reinforces them. If you want to infer process from pattern, be very very careful.
Incidentally, the “posture” of the beer cans explains the slope of the regression of Tb on Ta, but not why the histogram of Tb is left skewed. Some years later I puzzled over this and realized what is going on. Consider the body temperature of any inanimate object in the sun over a day. It starts cool, warms up, stabilizes at midday, and then cools in late afternoon. So the shape is somewhat of an upside and slightly squashed “U.” But if you extract the temperatures from that time series, you get a left skewed and right truncated histogram. Temperatures early and late are changing rapidly, so there are few of each temperature. Temperatures are stable at midday, and so there are lots of them. Thus, a skewed distribution is an expectation of a null model, and thus a skewed distribution for a real animal is not robust evidence of thermoregulation. However, this does not deny that thermoregulation could be occurring.
Let’s jump back to the botanist’s query, which in effect challenged my assumption that a U-shaped distribution of percentage of lizards in sun over the day implied thermoregulation. We found this pattern in all diurnal lizards we studied, and further showed that a higher percentage was in sun in winter than in summer – exactly what one would expect for a thermoregulator.
As Eric and I wrote our paper on thermoregulation of Kalahari lizards, I returned to the botanist’s challenge. I decided to take a Heath-like (null model) approach, and ask, what would be the expected percentage of lizards in the sun if lizards were moving randomly with respect to sun and shade and thus not thermoregulating.
I immediately realized that the answer depended on where the lizard lived. Consider a lizard that foraged on the ground. Early and late in the day, shrubs and trees will cast long shadows, and so little of the ground surface will be in sun; but at midday, shadows will be much reduced, and most of the ground surface will be in sun. Thus the null expectation is that the percentage of randomly moving lizards in sun will be low early and late in the day, and maximal at midday. Moreover, the percentage in sun at any time of day will be reduced in winter than in summer because the sun will be lower on the horizon, casting long shadows. However, our field data showed that lizards were doing just the opposite of the null, suggesting they were in fact thermoregulating (or perhaps tracking insects that were thermoregulating)!
Next consider a lizard on a tree trunk. Early and late in the day, up to 50% of the tree trunk is in sun, but at midday, little or none will be in sun, because tree’s crown will shade the trunk. That shading will be less at midday in winter, because the sun is lower on the horizon. In this case, the real lizards on a tree trunk are doing approximately the same as the null expectation, so in this case the observed pattern is not convincing of a process explanation of thermoregulation. Even so, non-random movement patterns would be implied if more than than 50% of the lizards are in sun early and late in the day (50% is the maximum theoretically possible for a non-regulator, because up to half of a regular tree trunk will always be in shade). In fact, the percentage of lizards we found in sun early and late in the day or in winter often exceeded 50%, consistent with a thermoregulation interpretation.
For our paper I wanted to draw some graphs of the expectations of sun use from a null model and asked Jeff Hoffman for help. Jeff was a roommate at Prentiss Street (Chapter 27) in Cambridge, a brilliant astrophysicist, and later an astronaut on five Shuttle missions. He knew navigation and trigonometry and was able to derive a model of sun and shade patterns, and we included his model as an appendix in our paper.
Some years later, Paul Hertz, Rob Stevenson, and I expanded our interest in null models and thermoregulation. We developed a method that used observed body temperature, operative temperatures (i.e., equilibrium temperatures on null lizards), and thermal preference to evaluate thermoregulation in the field.
Paul, Barry Sinervo, and I later used null models to show that behavioral thermoregulation can buffer selection on thermal sensitivity – a phenomenon we called “The Bogert Effect.” Our goal was to challenge the classic idea that behavioral shifts drive evolution. Martha Muñoz has greatly extended the evolutionary relevance of the Bogert Effect.
These models all trace to my chance encounter with a botanist in the Kalahari and to Jim Heath’s brilliant use of beer cans as null models. I appreciate that it is fun to ask, what is the pattern expected for an animal that is doing something like thermoregulation? However, it is even more fun to ask, what is the pattern expected for an animal that is not doing that thing but just moving randomly?
Jim was a creative scientist and had a sense of humor. Some examples are described an obituary that Dick Tracy and I wrote in Jim’s honor. I never knew him well, but he was one of the most influential people in my career.
For most people, thinking about ‘null’ patterns is unnatural. I had to work to think about null expectations. But learning to appreciate that concept is among the most useful concepts I have learned. Were Schlitz beer available in Seattle, I’d pop open a can and drink a toast to Jim Heath. He left us with a legacy.
I want to end with an example from real life – The Mystery of the Disappearing Forks. In the late ’70s and early ’80s, Joe Felsenstein, Monty Slatkin, and I usually ate lunch together. As we left the cafeteria, we would put our trays on conveyor belt. One day we saw a conspicuous sign above the belt asking people to turn in forks that they had taken to their offices and had failed to return to the cafeteria. Apparently a disproportionate number of forks were missing.
Monty pointed out that forks should be “lost” more frequently than knives or spoons, and that is is unlikely that people were selectively failing to return only forks. Why? As Monty noted, forks are the most generally useful utensil, and many people would pick up only a fork for their meal. Consequently, more forks should “disappear” from the cafeteria by chance and because people were preferentially failing to return forks. Monty’s null model for fork losses was a special case, but null models should be a central component of our educational system.