29 Janzen’s hypothesis
The major pinball event for my academic life occurred in January 1966. I’d just graduated from UC Berkeley and was taking a field course in tropical ecology in Costa Rica (Organization for Tropical Studies). I was still expecting to attend medical school the following September but was considering grad school in biology. In either case I needed to stay in school to avoid being drafted (Chapter 12). I’d never taken a formal course in ecology as an undergrad, and thus I arrived in Costa Rica knowing nothing about the tropics and little about ecology. But by the end of that course, I was leaning toward a career in biology.
Our first field site was at Finca Taboga, in Guanacaste Province (NW Costa Rica). We spent mornings exploring the local habitats, learning to identify some plants and animals, and doing simple field exercises. We spent our afternoons in plant and animal labs (learning to identify diverse organism) and listening to topical lectures.
Dan Janzen was one of the course personnel and led our first-morning walk. I was overwhelmed by the diversity and grandeur of the plants and animals Dam identified for us. Near the end of the morning, we came to a small pond and flushed a basilisk lizard, which fled by running across the pond. I had no idea that such adept lizards existed and blurted out, “Jesus Christ, look at that!” Janzen responded with a chuckle, “That’s what they’re called, ‘Jesus Christ lizards.’”
On one of the afternoons in Guanacaste, Dan gave a talk on “Why mountains were higher in the tropics.” [Note: when Dan published a paper on that theme in 1967 (in The American Naturalist), he had changed the title to “Why mountain passes are higher in the tropics). He argued that mountain passes are more effective barriers to dispersal in the tropics than in temperate regions. His argument is based on two premises: 1) that low- and high-altitude sites in the tropics have little overlap in thermal regimes and 2) tropical species should be relatively specialized for temperature. Consequently, a tropical pass will be more a physiological barrier than a pass in the temperature zone – hence, mountain passes are”higher” in the tropcis.
Dan’s lecture was my first introduction to conceptual ecology. It deftly blended multiple fields (climatology, thermal physiology, geography, distributions, dispersal, mountains, latitude, speciation, etc.). I was fascinated but never thought about doing research along these lines. But I did like Dan’s idea.
His concept festered in my brain until spring 1975, when I was trying to finish my Ph.D. thesis. I was working in my apartment and chanced upon a 1967 paper by Ron Heyer about distributions of herps along an altitudinal transect on Tilarán in Costa Rica. Heyer noted that the altitudinal ranges of herp species there were narrower than those of herps on a transect in Yosemite (Grinnell). Lights flashed. This pattern was a corollary on Janzen’s hypothesis! I immediately wanted to see if these patterns held for other mountain transects. I dropped my thesis and dashed to the library at the Museum of Comparative Zoology.
I spent the next week excitedly searching for monographs and articles that reported altitudinal distributions of herps. I extracted altitudinal range limits for each species and then computed an index of faunal turnover with elevation. [If Janzen’s hypothesis held, then tropical species should turn over relatively quickly with any given change in altitude.]
I compiled ten altitudinal transects from Tilarán (Costa Rica) to Lassen Peak (California). The pattern Heyer had reported held beautifully. That was among the most exciting weeks of my academic career. In a few days of intense library work, I had a new chapter for my thesis and the essence of a paper, though I didn’t publish it until 1978.
Hearing Dan’s lecture was a chance event. Looking back, I realize how much that lecture and his subsequent paper have ‘haunted’ my career – always in positive ways. It primed my interest in thermal physiology and ecology.
For most ecologists, the primary significance of Dan’s idea traces to its implications for tropical species diversity (enhanced isolation promotes speciation). But for me, here is a crucial sentence from Dan’s paper:
“…it is the temperature gradient across a mountain range which determines its effectiveness as a barrier, rather than the absolute height.”
As a hiker, I sometimes disagree with Dan’s dismissing of “absolute height”! Even so, Dan’s sentence resonated and caused me to wonder how temperature affects the performance and distribution of ectotherms. Why do environmental temperature gradients matter?
That thought train eventually led me to the idea of measuring organismal performance (e.g., speed, stamina, reproduction) at different body temperatures (now universally called “thermal performance curves”). Then, if one knew how an ectotherm’s body temperatures changed along an altitudinal transect (or between two habitats or two seasons), and if one also knew how temperature influenced that animal’s performance, then one could predict how performance would change along the transect (or between habitats or seasons). This was a simple but powerful idea.
For me, Dan’s theme serves as a conceptual mapping of some environmental variable onto performance. This concept can be generalized far beyond temperature. One variation popped up when I worked with Peter Ward on the impact of changing oxygen levels on maximum altitudinal distributions of animals living throughout the Phanerozoic. Using physiologically based simulations, we argued that geological periods of low oxygen would have limited animals to low elevations. Consequently, Pangea could not have been a “superhighway,” as commonly assumed. To this day I regret not titling the paper on paleo-distributions, “Why mountain passes were higher in the Triassic.”
Similarly, we used a simulation approach to explore how high hypothetical “paleo-mountaineers.” could have climbed over geological time (Huey and Ward, 2005b). Our simulations suggested that the low oxygen level in the Triassic would have prevented hypothetical “paleo-mountaineers” to reach Base Camp on Mt. Everest. This is all just a variation on a theme of Dan Janzen.
Let’s skip back a few decades. In winter 1977, I interviewed for an Assistant Professor position in the Department of Zoology at the University of Washington. I gave two seminars illustrating my “best stuff.” The ecology faculty was world class, and I was lucky to get the job. I joined the faculty the following September.
Shortly after I arrived, Don Farner (Chair of Zoology) told me he’d scheduled me for a departmental seminar in early October. I reminded him that I’d given two seminars earlier that year and had little new to say. Farner looked back at me, and I immediately started thinking about what I would cover in a third seminar. [Note: Farner had very bushy eyebrows and a powerful stare over his pipe. The look he gave me made deans melt.]
I titled my October seminar something like “A herpetological miscellany.” I talked about three short and unrelated topics. One was on Janzen’s hypothesis, and I showed that faunal turnover with altitude proceeded relatively rapid in the tropics.
While preparing my talk, I wondered, where are the highest mountain passes (that is, in absolute heights)? I knew that the major Andean and Himalayan peaks were all outside the tropics. Thus, if mountain passes were proportional to the absolute height of mountain peaks, then mountains and their passes must be higher in the temperate zone.
This tangent amused me. I scrounged data on peak heights and drew a figure (which I approximated below). So after I presented the real distributional data (soon published in The American Naturalist), I showed these new histograms, and I thus “corrected” Janzen – all in fun, of course.
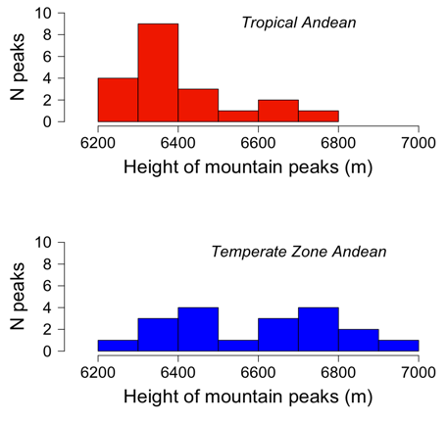
Next, I took the graph (which I had carefully inked by hand, mounted it on museum stock, and shipped it to Dan.
Almost two years elapsed before Dan responded. I saved his letter.

When that letter arrived, I was both stunned and amused, but wondered, would Dan really publish my graph in Biotropica? Every time I visited the UW library, at least for a year or so, I’d check the latest issue of Biotropica to see whether Dan had published my graph. He got back at me! And I had a good laugh at myself.
Janzen’s hypothesis continued to be in my life. Rickie van Berkum, one of my first grad students, did her thesis work, testing and supporting some assumptions underlying Janzen’s hypothesis (van Berkum 1976, 1978), specifically that tropical species were thermal specialists. Joyce Tsuji, another early grad student, another of Janzen’s assumptions, namely that the acclimation capacities of topical species was limited. All these studies were among the first to test Janzen’s hypothesis. .
Decades later, Cameron Ghalambor approached me at a meeting and asked, “Are you still interested in Janzen’s Hypothesis?” Of course! Cam suggested that we take a retrospective look at Janzen’s Hypothesis. I thought Cam’s idea was inspired, and we wrote that review (Ghalambor et al., 2006). Finally, to celebrate the 50th anniversary of Janzen’s paper in 2018, Kimberly Sheldon, Mike Kaspari, Nate Sanders, and I published a historical review of the impact of Janzen’s paper.
I would be remiss not to mention a related paper by Polato et al. (2018). This study combines field, physiological, and molecular approaches to test some implications of Janzen’s hypothesis. I was delighted to see this paper, which finally got to the core of Dan’s ideas. I was especially pleased that Kelly Zamudio, my last “herp” student, was a co-author.
Occasionally, I wonder what my career would have been like if I’d never heard (or paid attention to) Dan’s lecture and never read Ron Heyer’s paper. It would undoubtedly have been very different. I hope that I would have found something equally inspiring. But I don’t think I’d trade my “Janzen moment” (which I experienced in in the first few days of graduate school!) and the directions it led me to follow throughout my career.